viernes, 18 de diciembre de 2015
{kind=link}
martes, 20 de octubre de 2015
martes, 13 de octubre de 2015
sábado, 12 de septiembre de 2015
martes, 1 de septiembre de 2015
miércoles, 19 de agosto de 2015
domingo, 17 de mayo de 2015
miércoles, 22 de abril de 2015
lunes, 9 de marzo de 2015
lunes, 23 de febrero de 2015
martes, 17 de febrero de 2015
Anabolismo
TEMA
11: METABOLISMO I I. EL ANABOLISMO
- CONCEPTO DE ANABOLISMO
- ANABOLISMO HETERÓTROFO
- ANABOLISMO AUTÓTROFO
- FOTOSÍNTESIS
- FASE LUMINOSA
- FASE OSCURA
- BALANCE ENERGÉTICO DE LA FOTOSÍNTESIS
- FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS
- QUIMIOSÍNTESIS
- TIPOS DE ORGANISMOS QUIMIOSINTÉTICOS
1.
CONCEPTO DE ANABOLISMO
Conjunto
de procesos constructivos que suceden en una célula en los que se
produce la síntesis de moléculas complejas a partir de otras más
sencillas. El anabolismo es endergónico, necesita la incorporación
de energía.
Podemos
distinguir dos tipos de anabolismo:
Anabolismo
autótrofo.- Consiste
en la síntesis de moléculas orgánicas sencillas a partir de
precursores inorgánicos tales como el CO2,
el H2O
y el NH3.
Solamente pueden realizarlo las células autótrofas. Existen dos
modalidades de anabolismo autótrofo: la
fotosíntesis,
que utiliza la energía de la luz (en las células fotolitotrofas), y
la quimiosíntesis,
que utiliza la energía liberada en reacciones redox (el las células
quimiolitótrofas)
Anabolismo
heterótrofo.-
Consiste en la síntesis de moléculas orgánicas progresivamente más
complejas a partir de moléculas orgánicas más sencillas. Todas las
células pueden llevarlo a cabo (también las autótrofas). Utiliza
la energía del ATP y coenzimas reducidos que se obtienen en el
catabolismo.
2.-
ANABOLISMO HETERÓTROFO
Comprende
una serie de rutas anabólicas comunes (presentes en cualquier tipo
de célula). En
ellas se sintetizan macromoléculas fuertemente reducidas a partir de
moléculas orgánicas relativamente oxidadas.
Las
células autótrofas obtienen estas moléculas precursoras
fabricándolas en el anabolismo autótrofo, mientras que las células
heterótrofas las obtienen a partir del catabolismo de distintos
tipos de biomoléculas ingeridos en el alimento.
El
anabolismo heterótrofo es un proceso endergónico y como tal consume
energía química que es aportada por el ATP
y coenzimas reducidos. La mayoría de las reacciones del anabolismo
heterótrofo tienen lugar en el hialoplasma de la célula; algunas
finalizan en el retículo endoplasmático o en el aparato de Golgi.
Muchas
rutas del anabolismo heterótrofo recorren en parte el camino inverso
de las correspondientes rutas catabólicas aprovechando aquellas
reacciones que son claramente reversibles. Las reacciones
irreversibles se evitan dando "rodeos metabólicos".
Ejemplos
de procesos anabólicos en células heterótrofas:
Gluconeogénesis:Síntesis
de la glucosa a partir del ácido pirúvico. Esta ruta recorre en
gran parte el camino inverso de la glucólisis.
Glucogenogénesis:
El glucógeno se sintetiza en el hígado y en el músculo esquelético
con el objetivo de almacenar glucosa.se lleva a cabo a partir de
glucosa fosforilada en un proceso enzimático que consume energía
del UTP
o del ATP.
3.-
ANABOLISMO AUTÓTROFO
Las
rutas de síntesis de materia orgánica sencilla a partir de materia
inorgánica sólo son factibles en organismos autótrofos y se
diferencian en función de la fuente de energía que se utilice.
- Los organismos fotosintéticos utilizan la energía de la luz solar. Entre ellos están las plantas, las algas y algunas bacterias.
- Los organismos quimiosintéticos utilizan la energía liberada en las reacciones exergónicas. Son quimiosintéticas algunas bacterias como las del nitrógeno, del azufre o del hierro.
3.1.
LA FOTOSÍNTESIS
La
fotosíntesis es un proceso que llevan a cabo las células
fotolitótrofas
en el que, utilizando la energía luminosa capturada por ciertos
pigmentos, se sintetiza materia orgánica a partir de materia
inorgánica. Su ecuación global puede escribirse como sigue:
CO2
+ H2O
+ LUZ → MATERIA ORGÁNICA + O2
El
CO2
puede sustituirse en esta ecuación por sales minerales como nitratos
o sulfatos, que también se incorporan a la materia orgánica por
este procedimiento.
La
fotosíntesis tiene lugar principalmente en los cloroplastos,
siendo la principal función que desempeña este orgánulo en las
células de las plantas verdes y de las algas. Sin embargo, algunas
células procariotas (como ciertas bacterias y las algas cianofíceas)
también realizan la fotosíntesis a pesar de no poseer estos
orgánulos, ya que poseen pigmentos fotosintéticos asociados a sus
respectivas membranas plasmáticas.
Las
reacciones de la fotosíntesis pueden agruparse en dos grandes
bloques:
- la fase luminosa, en la que la energía de la luz capturada por los pigmentos fotosintéticos se transforma en energía química del ATP y NADPH.
- la fase oscura, en la que la energía acumulada en estos dos compuestos es utilizada para transformar el dióxido de carbono y las sales minerales en materia orgánica.
La
membrana tilacoidal de los cloroplastos, además de los lípidos y
las proteínas característicos de toda membrana, posee un 12% de
otras sustancias que se denominan, debido a su capacidad para
absorber luz, pigmentos
fotosintéticos.
Existen dos tipos principales de pigmentos: las clorofilas
y
los carotenoides.
Ambos tienen en común el poseer un sistema de dobles enlaces
conjugados (dobles enlaces que se alternan con enlaces sencillos).
Esta circunstancia es la que les permite absorber la energía
luminosa, ya que los electrones de éste sistema de dobles enlaces
conjugados pueden "excitarse", es decir, pasar a niveles
energéticos superiores, sin que la molécula se rompa. Los distintos
tipos de pigmentos están especializados en absorber luz de una
determinada longitud de onda, por lo que, actuando de modo
coordinado, cubren todo el espectro de la luz visible.
Los
pigmentos fotosintéticos no están distribuidos al azar por la
membrana tilacoidal sino que están organizados formando
fotosistemas.
Un
fotosistema es
una unidad funcional constituida por:
a)Un
complejo antena.-
Está formado por varios centenares de moléculas de clorofilas,
carotenoides y también proteínas, que absorben la energía de la
luz de diferentes longitudes de onda y la canalizan hacia una única
molécula de clorofila denominada clorofila
diana.
La antena funciona como una especie de embudo para capturar la
energía luminosa
b)Un
centro de reacción.-
Está formado por la clorofila diana, un dador de electrones y un
aceptor de electrones que pueden variar de unos fotosistemas a otros.

Existen
dos tipos de fotosistemas: el fotosistema
I (PS I o P700) y
el fotosistema
II (PS II o P680),
que difieren en el tipo de clorofila diana que poseen.
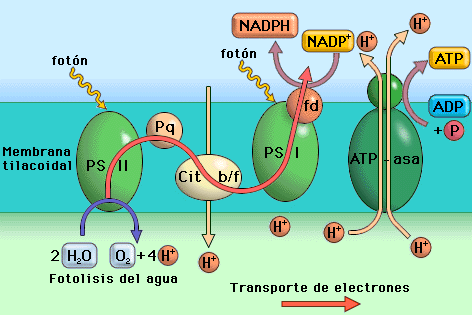
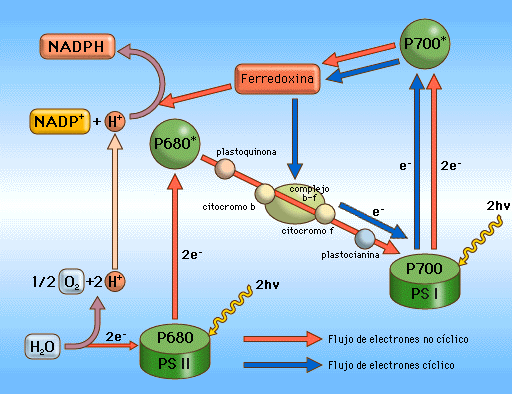
*
FASE LUMINOSA
Consiste
en un transporte
de electrones
a través de una cadena transportadora ubicada en la membrana
tilacoidal de los cloroplastos. Mientras que en la mitocondria el
transporte electrónico se realizaba a favor de gradiente de
potencial redox, es decir, desde buenos dadores de electrones a
buenos aceptores, en el cloroplasto este transporte se realiza en
sentido contrario,
desde el H2O,
que es un débil dador de electrones, hasta
el NADP+,
que es un débil aceptor. Este transporte electrónico "cuesta
arriba" es un proceso
endergónico,
y no tendría lugar si no se le suministra energía. Aquí es donde
interviene
la energía luminosa
captada por los pigmentos fotosintéticos: es utilizada para impulsar
los electrones desde el agua hasta el NADP+,
que se reduce entonces para dar NADPH.

1.- El proceso comienza cuando la luz excita un par de electrones del PS II, que se oxida, y puede aceptar ahora otros dos electrones procedentes del agua volviendo a su estado inicial. Para ello la molécula de agua se rompe (fotolisis del agua) liberando así el oxígeno, que es uno de los productos de la fotosíntesis.
2.-
Los electrones excitados del PS II son cedidos a la cadena de
transportadores.
3.-
Al mismo tiempo que esto sucedía, un par de electrones del PS I
fueron excitados por la luz y cedidos a otro tramo de la cadena que
los conduce al NADP+
que se reduce a NADPH.
Los electrones procedentes del PS II son cedidos ahora al PS I que
recupera su estado inicial.
Fotofosforilación
no cíclica
Aunque
globalmente el transporte electrónico fotosintético se realiza
"cuesta arriba", el uso combinado de los dos fotosistemas
permite la existencia de un tramo descendente en el que los
electrones circulan a favor de gradiente de potencial redox. En este
tramo se libera algo de energía que, de modo similar a lo que ocurre
en la mitocondria, es utilizada para bombear protones a través de la
membrana tilacoidal (desde el estroma al espacio intratilacoide),
creando así un gradiente electroquímico de protones. En esta
membrana también existe una ATP-sintetasa
que, gracias a la energía liberada al regresar los protones al
estroma, puede sintetizar ATP.
Dado
que el tramo descendente al que aludimos es corto comparado con la
totalidad de la cadena, se bombean muy pocos protones y sólo
se obtienen una molécula de ATP
por cada par de electrones que circulan desde el agua al NADP+.
Fotofosforilación
cíclica
Haciendo
un balance de lo ocurrido se observa que por cada molécula de agua
que se rompe por fotolisis se obtiene una
molécula de NADPH
y una molécula de ATP.
Esto plantea un problema, ya que, como veremos, en la fase oscura se
necesita más cantidad de ATP
que de NADPH.
Por ello, las células fotosintéticas se ven obligadas a recurrir a
un mecanismo alternativo al que hemos descrito que les permita
obtener ATP
sin obtener NADPH.

Este
mecanismo alternativo recibe el nombre de fotofosforilación
cíclica
y consiste en lo siguiente: los electrones excitados del PS I, en
lugar de ser cedidos al NADP+,
lo son a los transportadores del tramo descendente de la cadena,
regresando al PS I; en este regreso se libera energía suficiente
para bombear protones y obtener ATP.
A diferencia de lo que ocurre en la fotofosforilación no cíclica,
no participa el agua, no se libera oxígeno, no participa el PS II y
no se obtiene NADPH.
FASE
OSCURA
Consiste
en la síntesis de moléculas orgánicas sencillas por reducción de
moléculas inorgánicas, utilizando el NADPH y el ATP obtenidos en la
fase luminosa. Se produce en el estroma del cloroplasto y puede
suceder en presencia o en ausencia de luz.
El
principal sustrato utilizado es el CO2
que es reducido a monosacáridos sencillos, pero también es posible
la utilización de nitratos o sulfatos.
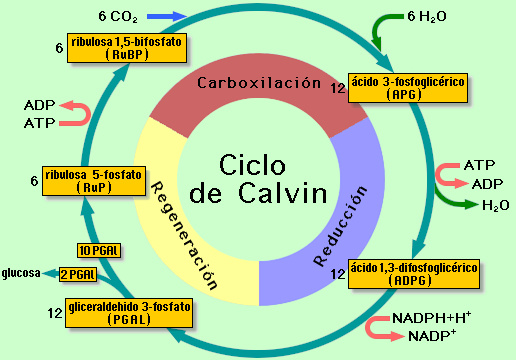
LA
FIJACIÓN DEL CO2:
EL CICLO DE CALVIN
- Fijación del CO2: El CO2 se fija inicialmente sobre la ribulosa-1,5-difosfato (5C), que es la molécula aceptora, dando lugar a 2 moléculas de ácido fosfoglicérico (3C). Esta reacción está catalizada por la enzima ribulosa 1,5-difosfato carboxilasa o rubisco, que es la enzima más abundante de la naturaleza y tiene la peculiaridad de ser relativamente lenta, por lo que constituye un verdadero factor limitante para la velocidad de la fotosíntesis. Además este enzima puede utilizar como sustrato al oxígeno en lugar del dióxido de carbono, dando lugar a una secuencia de reacciones que terminan liberando CO2. Este fenómeno se conoce como fotorrespiración y es responsable de una sensible disminución en el rendimiento de la fotosíntesis (la fotosíntesis fija CO2 y la fotorrespiración lo libera). Algunas plantas tropicales, llamadas plantas C4, evitan este problema fijando inicialmente el CO2 mediante una ruta alternativa al ciclo de Calvin, la ruta de Hatch y Slack, en la que el enzima responsable de la fijación no se ve interferido por el oxígeno.
- Reducción: El ácido fosfoglicérico, a expensas del ATP y NADPH procedentes de la fase luminosa, se reduce a gliceraldehido-3-fosfato, que está en equilibrio con su isómero, la dihidroxiacetona-fosfato.
- Recuperación y síntesis de biomoléculas: Estos dos fosfatos de triosa son utilizados en parte para regenerar la ribulosa-difosfato mediante una serie de reacciones que implican gasto de ATP, y en parte son desviados hacia el anabolismo heterótrofo para servir de precursores a distintos tipos de biomoléculas.
Cada
vuelta del ciclo de Calvin implica la fijación de 1 CO2 (=1C), por
tanto, para generar una molécula de glucosa (6C) se necesitan 6
vueltas.
En
este caso, la ecuación global del ciclo sería:
6CO2
+ 12 NADPH + 12H+
+ 18 ATP → C6H12O6
+ 12 NADP+
+ 18 ADP + 18 Pi
BALANCE
ENERGÉTICO DE LA FOTOSÍNTESIS:
La
fase luminosa de la fotosíntesis produce ATP y NADPH. Si se
sintetiza una molécula de glucosa (C6H12O6) se necesitan 6 CO2 y 12
de Agua. El agua libera 6 O2 a la atmósfera y aporta 12 hidrógenos
de la glucosa y los 12 hidrógenos necesarios para pasar los 6 O2
sobrantes del CO2 a Agua. Intervienen 24 Hidrógenos. Aparecen así
24 protones y 24 electrones y, como cada electrón precisa dos
fotones (uno en el PSI y otro en el PSII), se necesitan 48 fotones.
El ciclo de Calvin necesita por cada CO2 incorporado, 2 NADPH y 3
ATP. Para una molécula de glucosa se necesitan 12 NADPH y 18 ATP.
FACTORES
QUE INFLUYEN EN LA FOTOSÍNTESIS
Dado
que la fotosíntesis es un proceso que consume CO2
y desprende O2,
la intensidad
fotosintética puede
determinarse experimentalmente midiendo el volumen de CO2
consumido o de O2
desprendido.
Los
factores ambientales que afectan a la fotosíntesis son:
Concentración
de CO2
en el medio.-
Si la iluminación es constante, la intensidad fotosintética
aumenta con la concentración de dióxido de carbono en el medio.
Esto es debido a que a mayor concentración de CO2
mayor será la velocidad a la que la ribulosa difosfato carboxilasa
fijará el CO2
en el ciclo de Calvin.
Intensidad
de la iluminación.-En
general la intensidad de la iluminación aumenta el rendimiento de
la fotosíntesis. Esto es debido a que cuantos más fotones de luz
lleguen a los fotosistemas I y II, más cantidad de ATP
y NADPH
se obtendrán en la fase luminosa y en consecuencia más CO2
se podrá fijar en la fase oscura. Sin embargo, si se sobrepasa un
cierto límite de intensidad luminosa puede producirse la
desnaturalización de los enzimas que actúan en el proceso con lo
que la intensidad fotosintética disminuye bruscamente.
Si
la concentración de CO2
es muy baja la iluminación apenas influye sobre la intensidad
fotosintética (de poco vale que se obtenga mucho ATP
y NADPH
en la fase luminosa si no hay CO2
para fijar en la fase oscura). Sin embargo, a partir de un cierto
valor en la concentración de CO2
la intensidad de iluminación se convierte en un factor limitante
para el proceso fotosintético.
Concentración
de oxígeno.-
El O2
tiene un efecto inhibitorio sobre la fotosíntesis: a mayor
concentración de O2
en el medio menor es la intensidad fotosintética. Esto es debido al
fenómeno de la fotorrespiración (Véase la actividad de la enzima
rubisco).
Temperatura.-
Las reacciones fotoquímicas de la fase luminosa son independientes
de la temperatura. Sin embargo, las reacciones de la fase oscura
incrementan su velocidad con la temperatura tal como ocurre en
cualquier reacción química. Todo ello siempre que no se alcance un
valor tal que se provoque la desnaturalización de los enzimas, en
cuyo caso la intensidad fotosintética disminuye bruscamente.
Humedad.-
El rendimiento de la fotosíntesis disminuye en ambientes áridos
porque al cerrarse los estomas para evitar la pérdida de agua
también se reduce el intercambio de gases.
Color
de la luz.- El máximo
rendimiento de la fotosíntesis se obtiene con luz roja o azul. Una
luz con una longitud de onda superior a los 680 nm impide la
actuación del fotosistema II y, por tanto, sólo se produce
fotofosforilación cíclica con un rendimiento menor.
3.2.
LA QUIMIOSÍNTESIS
La
quimiosíntesis es un proceso que llevan a cabo las células
quimiolitótrofas, las cuales son capaces de utilizar la energía
liberada en la oxidación de moléculas inorgánicas sencillas para
fijar la materia inorgánica en forma de materia orgánica. Sólo un
reducido número de bacterias, entre las que destacan las bacterias
del suelo que oxidan el amoníaco a nitritos y estos a nitratos,
pueden llevar a cabo este proceso.
La
quimiosíntesis, de manera análoga a la fotosíntesis consta de dos
fases:
a)
En la primera
fase,
análoga a la fase luminosa de la fotosíntesis, se obtiene energía
química en forma de ATP
y coenzimas reducidos a partir de la oxidación de compuestos
inorgánicos sencillos tales como amoníaco, nitritos, azufre y sus
derivados, hierro y otros. Cada grupo de organismos quimiosintéticos
está especializado en la obtención de energía a partir de un
determinado tipo de compuestos inorgánicos.
b)
En la segunda
fase,
el ATP
y coenzimas reducidos obtenidos en la fase anterior son utilizados
para reducir compuestos inorgánicos transformándolos en compuestos
orgánicos. Esta segunda fase es muy similar, tanto para el carbono
como para el nitrógeno, a la fase oscura de la fotosíntesis.
Los
organismos quimiosintéticos presentan una serie de características
comunes:
- Son procariotas autótrofas. Solamente algunas bacterias poseen metabolismo quimiosintético.
- Viven de una fuente inorgánica: agua, sales, O2, CO2 y compuestos inorgánicos de cuya oxidación obtienen energía.
- Obtienen la energía de una reacción química específica. Solamente crecen con compuestos específicos de origen inorgánico, o producidos por la actividad de otros organismos (descomposición, excreción).
- Son aerobios. Utilizan el oxígeno como último aceptor de electrones.
- Sintetizan materia orgánica por medio del ciclo de Calvin.
Los
microorganismos quimiosintéticos cierran los ciclos biogeoquímicos,
posibilitando la vida en el planeta y devolviendo al sustrato las
sustancias procedentes de la oxidación de materia de descomposición
de los organismos muertos. De este modo, los restos de los seres
vivos se transforman en sales minerales de nitrógeno o azufre
(mineralización de la materia orgánica) que pueden ser de nuevo
absorbidas por los vegetales.
TIPOS
DE ORGANISMOS QUIMIOSINTÉTICOS
- BACTERIAS DEL NITRÓGENO : Viven en el suelo y en el agua. Utilizan como sustratos compuestos reducidos de nitrógeno. Oxidan el amoníaco procedente de la descomposición de la m.o. a nitratos (nitrificación). Este proceso se produce en dos fases en las que intervienen dos grupos de bacterias diferentes:
- Bacterias nitrosificantes: Oxidan el amoníaco hasta nitritos (G. Nitrosomonas).
- Bacterias nitrificantes: Oxidan los nitritos hasta nitratos (G. Nitrobacter)
- BACTERIAS INCOLORAS DEL AZUFRE: Viven en aguas residuales, fuentes hidrotermales y ambientes ricos en azufre o sus derivados. Utilizan como sustrato azufre y sulfuro de hidrógeno.
- BACTERIAS DEL HIERRO O FERROBACTERIAS: Viven en aguas procedentes de vertidos mineros. Oxidan las sales ferrosas y obtienen sales férricas.
- BACTERIAS DEL HIDRÓGENO: Utilizan el hidrógeno como sustrato. En su mayoría son quimioautótrofas facultativas.
jueves, 8 de enero de 2015
Suscribirse a:
Comentarios (Atom)